| Issue |
Ann. For. Sci.
Volume 67, Number 4, June 2010
|
|
|---|---|---|
| Article Number | 411 | |
| Number of page(s) | 12 | |
| DOI | https://doi.org/10.1051/forest/2009130 | |
| Published online | 02 April 2010 | |
Original article
The importance of canopy-derived dissolved and particulate organic matter (DOM and POM) – comparing throughfall solutionfrom broadleaved and coniferous forests
L’importance des dérivés dissous et des particules de matière organique (DOMet POM) de la canopée – comparaison des solutions des précipitations par égouttement sous les couverts des forêts feuillues et des conifères
1
Geographical Institute, Section Landscape Ecology,
Goldschmidtstrasse
5, 37075
Göttingen, Germany
2
NW-FVA. Northwest German Forest Research Station, Department Environmental
Control, Grätzelstr. 2, 37077
Göttingen, Germany
3
Friedrich-Schiller-University of Jena, Institute of Geography, Chair in Soil Science
Löbdergraben
32, 07743
Jena, Germany
* Corresponding author:
This email address is being protected from spambots. You need JavaScript enabled to view it.
Received:
28
April
2009
Accepted:
21
September
2009
Abstract
• Dissolved organic matter (DOM) and its main constituents carbon (DOC) and nitrogen (DON) represent an important part of the C and N cycles in forest ecosystems. Although many investigations have been addressing this issue, the knowledge on particulate organic matter (0.45 μm < POM < 500 μm) dynamics, its origin and involvement in organic matter cycling in forest ecosystems is still imperfect.
• In this paper, we report on dissolved and particulate organic carbon and nitrogen fractions in throughfall solutions collected from a broadleaved and coniferous forest stand in Central Germany. Over a period of 2.5 y (2005–2007) we followed the concentrations and fluxes of DOM and POM at a mature beech (Fagus sylvatica L.) and a Norway spruce (Picea abies L.) forest site. Bulk and throughfall precipitation were sampled in weekly (2005) and fortnightly (2006–2007) intervals and analyzed for dissolved (< 0.45 μm, filtered) and total (< 500 μm, unfiltered) amounts of organic carbon (DOC, TOC, POC) and nitrogen (TN, DN, PON, NO3-N) species. Proportions of particulate organic C and N were determined by difference between total and dissolved fractions.
• Under spruce, throughfall concentrations of most C and N fractions were twice as high as under beech. At both sites, concentrations and fluxes were significantly higher during the growing than the dormant season. At the broadleaved site, 80% of the annual fluxes of the DOC and TOC and 70% of the DN and TN were released during the growing season, compared to 60% for C and N at the coniferous site. POC under beech contributes with up to 30% to TOC compared to less than 20% at the spruce site.
• We suggest that pollen deposition, insect excretions and accumulated organic matter mobilised by dry/wet precipitation patterns play a supreme role for the formation of DOM and POM in forest canopies. The study demonstrates that the canopy is an important source for POM. Dynamics of DOM and POM are mainly driven by tree species effects and seasonality as well as by biotic agents.
Résumé
• La matière organique dissoute (DOM) et ses principaux constituants carbonés (DOC) et l’azote (DON) représentent une part importante des cycles du carbone et de l’azote dans les écosystèmes forestiers. Bien que de nombreuses enquêtes se soient penché sur ce problème, les connaissances sur les dynamiques des particules de matière organique (0,45 μm < POM < 500 μm), leur origine et leur rôle dans les cycles de la matière organique dans les écosystèmes forestiers sont encore imparfaites.
• Dans ce papier, nous présentons un rapport sur les particules de carbone organique et les fractions d’azote dissoutes dans les solutions de pluie arrivant directement au sol sous le couvert, récoltées dans un peuplement forestier feuillu et dans un peuplement de conifères en Allemagne centrale. Sur une période de 2,5 ans (2005–2007) nous avons suivi les concentrations et les flux de DOM et de POM dans une hêtraie arrivée à maturité (Fagus sylvatica L.) et un peuplement d’épicéa commun (Picea abies L.). Le volume des pluies et les précipitations au sol ont été échantillonnés à des intervalles de temps d’une semaine (2005) et d’une quinzaine (2006–2007) et analysés pour les quantités de carbone organique dissoutes (< 0,45 μm, filtrées) et totales (< 500 μm, non filtrées) (DOC, TOC, POC) et d’azote (TN, DN, PON, NO3-N). Les proportions des particules organiques de carbone et d’azote ont été déterminées par différence entre les fractions totales et dissoutes.
• Sous la pessière, les concentrations de la plupart des fractions de C et de N, dans la pluie au sol, étaient deux fois plus élevées que sous la hêtraie. Sur les deux sites, les concentrations et les flux ont été significativement plus élevés pendant la période de croissance que durant la période de dormance. Sur le site feuillu, 80 % des flux annuels de DOC et de TOC et 70 % de DN et TN ont été libérés au cours de la saison de croissance, contre 60 % pour C et N pour les conifères. Dans la hêtraie POC contribue à hauteur de 30 % de TOC, comparativement à moins de 20 % dans la pessière.
• Nous suggérons que les dépôts de pollen, les déjections d’insectes et la matière organique accumulée, mobilisés par les séquences de périodes sèches et de précipitations jouent un rôle suprême pour la formation des DOM et POM dans les canopées forestières. L’étude démontre que la canopée est une source importante pour POM. Les dynamiques de DOM et POM sont principalement conduites par les effets des espèces d’arbres et par la saisonnalité ainsi que par des agents biotiques.
Key words: dissolved organic matter (DOM) / particulate organic matter (POM) / throughfall / fluxes and concentrations / temperate forests
Mots clés : matière organique dissoute (DOM) / particules de matière organique (POM) / pluie au sol par égouttement / flux et concentrations / forêts tempérées
© INRA, EDP Sciences, 2010
1. INTRODUCTION
Fluctuations in throughfall composition have been reported from coniferous (Manderscheid and Matzner, 1995) and also from deciduous forests (Currie et al., 1996). Annual throughfall concentrations of dissolved organic carbon (DOC) in temperate forest ecosystems highly vary between 3 and 35 mgL−1, those of dissolved organic nitrogen (DON) between 0.25 and 1.11 mgL−1.
As principle drivers behind differences in throughfall composition, divers abiotic and biotic processes likely take part in forming varying energy and nutrient transfer patterns. In this context, studies demonstrated that precipitation patterns (allowing organic matter to accumulate in the canopy during dry periods and to get remobilized during re-wetting events) (Mercier and Lindow, 2000), canopy architecture (Levia and Frost, 2006), atmospheric ecosystem deposition (e.g. pollen, inorganic N deposition) (Lamersdorf and Blank, 1995), increased nutrient leaching from damaged leaves and immobilization processes within the canopy mediated by phyllosphere micro-organisms (Stadler and Müller, 2000) are considered to be responsible for different release patterns.
While several investigations have sought to understand the nature and dynamics of DOM in terrestrial ecosystems (Currie et al., 1996), only few studied the dynamics, origin and involvement of particulate organic matter (0.45 μm < POM < 500 μm) in nutrient cycling of forested ecosystems. We are only aware of three studies, which were conducted by Carlisle et al. (1966) in a mature oak stand, by Sollins et al. (1980) reporting on micro-particulate matter fluxes ( < 0.5 μm) with throughfall in an old-growth Douglas-fir forest ecosystem, and by Lamersdorf and Blank (1995) within the Solling roof project under spruce. The latter exhibited a significant additional input of organic carbon and nitrogen in throughfall via the particulate fraction amounting to 11–15% for carbon and to 14–21% for nitrogen relative to the total annual input by litter-fall and throughfall (< 0.45 μm). Additionally, the low C to N ratios between 12–14 supply evidence for an enhanced susceptibility to fast microbial decomposition. Recently, le Mellec and Michalzik (2008) found a significant contribution of particulate organic N (PON) providing an extra input of up to 30% to the dissolved N fluxes with throughfall in pine forest stands in northern Germany. Consequently, all studies demonstrated the quantitative importance of the particulate organic matter fraction and its consideration within the C and N budget of forests. However, detailed information on the nature, origin and the environmental factors governing the dynamics of canopy-derived POM are still scarce.
The transfer of organic matter and nutrients between tree canopies and the soil compartment is regarded as an important connective pathway between the above and below ground system, exhibiting e.g. inter-linkages between tree species and spatial and temporal patterns of soil acidity and cation cycling in temperate forests (Finzi et al., 1998). Additionally, field studies and meta-analyses of literature data support the idea, that canopy-derived organic matter inputs considerably affect organic matter and nutrient dynamics in the forest floor. For temperate forest ecosystems, Michalzik et al. (2001) found that variations in annual DOC and DON throughfall fluxes explain 46% and 65% of the variability in annual DOC and DON fluxes from the forest floor, while water fluxes alone did not explain a significant portion of the variability.
To reveal tree canopy-soil interactions the accurate determination of input fluxes to the soil is of central importance. However, since element and nutrient fluxes are conventionally measured after standard filtration (0.45 μm pore size), the exclusion of the particulate/unfiltered organic matter fraction (0.45 μm < POM < 500 μm) potentially results in misleading inferences and budgeting gaps when studying nutrient and energy fluxes in ecosystems.
Yet another aspect might come into focus, dealing with feedback mechanisms covering the climatic change-associated increase in atmospheric CO2 concentrations and effects on throughfall composition. In this context, Lichter et al. (2000) investigated in a FACE (Free Air CO2 Enrichment) study the impact of increased CO2 concentrations ( ∼ 560 ppm) on throughfall composition gathered in a loblolly pine forest. They found that within two years the DOC deposition increased by about 48% compared to the control, apparently indicating an enhanced availability of leaf-derived easy soluble C compounds. Findings by Hagedorn et al. (2008) for alpine forest ecosystems corroborate trends of increased DOC due to increased atmospheric carbon concentrations. According to these results, it is worthwhile to study the current transfer amounts of OM in solutions from the canopy to the soil to determine a basis for assessing future changes. Additionally, human-induced impacts (e.g. air pollution) lead to enhanced atmospherically depositions, especially of inorganic N-species, which affect input situations and thus the biogeochemistry of forest ecosystems, and plausibly canopy processes as well (Aber et al., 1989; Rennenberg and Gessler, 1999).
In the present study we aim to clarify the following hypotheses: (i) the canopy is a relevant source also for particulate organic matter (POM) within the C and N budget of forest ecosystems; (ii) the quantity and temporal dynamics of DOM and POM in broadleaved forests (beech) differ from those in coniferous ones (spruce); (iii) canopy-derived POM is built up by different sources, which encompass pollen, atmospheric deposits, leached plant biomass, phyllosphere microbial biomass and excretions of phytophagous insects and finally that (iv) different environmental factors contribute to the release of POM.
To test our hypotheses, we followed the concentrations and fluxes of DOM and POM at a mature beech (Fagus sylvatica L.) and a Norway spruce (Picea abies L.) forest site in Central Germany over a period of two and a half years (2005–2007). Bulk and throughfall precipitation were sampled in weekly (2005) and fortnightly (2006–2007) intervals and analyzed for dissolved ( < 0.45 μm, filtered) and total ( < 500 μm, unfiltered) amounts of organic carbon (DOC, TOC, POC) and nitrogen (TN, DN, PON, NO3-N) species. Proportions of particulate organic C and N were determined by difference between total and dissolved fractions.
2. MATERIALS AND METHODS
2.1. Site description
The study was carried out at the Level II monitoring sites B1 and F1, a mature beech and spruce stand, respectively, in the Solling mountain area (Lower Saxony, Germany) located at 500 m a.s.l. The climate is classified as humid and sub-continental with a low mean annual temperature (6.9 °C), and high mean values of precipitation (1193 mm) and relative humidity (82%) (period 1990 to 2002, Panferov et al., 2010).
The coniferous site (F1) is planted on former grassland with Norway spruce (Picea abies Karst) of about 125 y of age and is approximately 1 ha large. The stand density is 465 treesha−1 and the leaf area index (LAI) shows values of 20.5 m2m−2 to 33 m2m−2 (Ibrom, 1993). The growing stock (timber volume) amounts to 733 m3ha−1 and is classified as very dense (Meesenburg, unpublished). The understory vegetation forms patches of grass, moss and blueberry (Vaccinium myrtillus) vegetation.
The beech (Fagus sylvatica L.) site (B1) is approximately 160 y old and exhibits nearly vegetation-free ground conditions. The stand density counts 193–199 trees ha−1, the growing stock is 519 m3ha−1 and the LAI is 6.5 m2m−2 (Meesenburg and Brumme 2009; Schipka et al. 2005). Since the early nineties both sites form part of the Level II monitoring program of the UN/ECE ICP forests. Deposition monitoring is performed at both sites since 1968 (Matzner and Meiwes, 1994).
At both sites, soils have developed from the same parent material encompassing a loess layer (eolian silt cover) over Triassic sandstone, which is typical of the Solling region (Ellenberg et al., 1986).
2.2. Experimental set up
Our investigations covered a period of approximately 2.5 y, starting in mid-April 2005 ending in mid-December 2007. The analyses focused on the determination of organic C and N concentrations in throughfall and bulk solutions.
Bulk precipitation was collected in three replicates at an open area near the forest site using open funnel samplers of 326 cm2 sampling area. Throughfall solution was sampled by collectors of the same type, arranged in a grid scheme (3 rows with 5 samplers each), yielding 15 replicates each on the spruce and the beech site. Five throughfall samples each were pooled, resulting in 3 mixed throughfall samples per study plot and sampling date. Sampling was performed in weekly intervals from April to December 2005, and in fortnightly intervals for the rest of the sampling period. Element fluxes were calculated from measured water volumes and element concentrations.
In 2006 a significant part of the spruce site was infested by bark beetles requiring a partial clear cut of the study plot. Consequently, sampling at the spruce site was interrupted from August 8th to October 31st, until the rain samplers were moved to an adjacent area within the same stand 20 m away from the former one. In late September 2006, the beech plot was infested for a short time by phytophagous larvae of the lymantrid moth (Dasychira pudibunda), though without changing the sampling protocol.
2.3. Chemical analysis
In our study we considered following compounds and fractions: total organic carbon (TOC) which includes the fractions dissolved organic carbon (DOC < 0.45 μm) and particulate organic carbon (> 0.45 POC < 500 μm). Moreover, total nitrogen (TN) which consists of the fraction dissolved nitrogen (DN < 0.45 μm) and particulate nitrogen ( > 0.45 PN < 500 μm). DN includes the inorganic (NH4, NO3) and organic (DON) forms.
One aliquot of the bulk/throughfall solutions was 0.45 μm
membrane-filtered (Cellulose-acetate filters, Sartorius). The analysis included the
determination of dissolved organic carbon (DOC) and dissolved nitrogen (DN) by thermal
oxidation (Dimatoc 100, Dimatec, Essen, Germany), and NO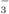 by ion chromatography
(761 Compact IC, Metrohm). From the difference between DN and NO3-N we
calculated residual-N consisting of NH4-N and dissolved organic nitrogen (DON).
by ion chromatography
(761 Compact IC, Metrohm). From the difference between DN and NO3-N we
calculated residual-N consisting of NH4-N and dissolved organic nitrogen (DON).
Average annual concentrations (mg L−1) of different C and N fractions in throughfall solutions (mean ± standard deviation) of the beech and spruce site at Solling (2005–2007).
In unfiltered aliquots, total organic carbon and total nitrogen (TOC and TN < 500 μm) were also assessed by thermal oxidation (Dimatoc 100, Dimatec, Essen, Germany), where the upper particle size limit of 500 μm was given operationally by the capillary diameter of the TOC/TN-analyzer. Particulate organic carbon (POC) was calculated as difference between TOC and DOC and particulate organic nitrogen (PON) as difference between TN and DN. Since the dissolved species nitrate and ammonium do not contribute to the particulate N fraction ( > 0.45 μm), we therefore hypothesize, that PON basically represents organically bound nitrogen (e.g. in proteins derived from cell fragments, and microbial biomass N. In addition, the solution pH (WTW, SenTix 61, glass electrode) and electrical conductivity (EC) (WTW, TetraCon 325) were determined in unfiltered solutions.
2.4. Statistical analysis
The non-normal distribution of the data implicated the application of non-parametric tests. Accordingly, we applied the Spearman’s test for correlation analysis and the Man-Whitney-test (U-test) to test for statistical differences. All tests were carried out by program packages of SPSS Statistics.
3. RESULTS
3.1. Concentrations in throughfall solutions
For most of the dissolved and total C and N species (with the exception of PON) average throughfall concentrations under spruce were significantly higher than those under beech (Tab. I). Statistical tests exhibited significant differences between the two tree species. Average throughfall concentrations under beech were 8.7, 1.9 and 10.8 mgL−1 for DOC, POC and TOC, respectively, and 2.79, 0.81, 0.80 and 3.14 mgL−1 for DN, PON, NO3-N and TN. With the exception of PON, corresponding values under spruce were approximately twice as high exhibiting 15.1, 2.9 and 18.0 mgL−1 for DOC, POC and TOC, respectively, and 5.23, 0.58, 1.85 and 8.20 mgL−1 for DN, PON, NO3-N and TN (Tab. I).
 |
Figure 1 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved and particulate organic C fractions in throughfall solutions of the beech stand (2005–2007). |
Independent from tree species, concentration levels were found to vary annually with minimum average concentrations occurring in 2007 (Tab. I). Furthermore, for DOC, TOC, DN and TN, throughfall concentrations exhibited pronounced differences between the growing and the dormant season with statistically significantly higher concentrations during the growing season at a P-level < 0.001 (Man-Whitney-U-test, data not shown).
3.2. Temporal dynamics of throughfall concentrations
At both sites, the concentrations of DOC, TOC, DN and TN were temporally highly fluctuating revealing a relation to seasonal patterns which showed larger values during the summer months compared to the rest of the year (Figs. 1–4). Dissolved N composition at both sites exhibited higher proportions of residual-N, representing DON and NH4-N, during the warmer season of the year (Figs. 5 and 6).
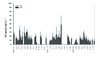 |
Figure 2 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved and particulate organic C fractions in throughfall solutions of the spruce stand (2005–2007). |
 |
Figure 3 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved and particulate N fractions in throughfall solutions of the beech stand (2005–2007). |
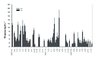 |
Figure 4 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved and particulate N fractions in throughfall solutions of the spruce stand (2005–2007). |
 |
Figure 5 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved N fractions in throughfall solutions of the beech stand (2005–2007). |
 |
Figure 6 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved N fractions in throughfall solutions of the spruce stand (2005–2007). |
At the beech site, single abiotic and biotic events such as September 19th 2005, May 22nd and September 9th 2006 and May 15th 2007 considerably magnified (up to 4–8 times) the concentrations of all C and N fractions especially the particulate ones (Figs. 1, 3 and 5).
The date in 2005 marked a period with high rain amounts which followed a one-month drought period in April, likely washing-off previously accumulated matter from the canopy. In May 2006 remarkable amounts of spruce pollen were released from the canopy, significantly enhancing POC and PON under beech. In September 2006 phytophagous larvae attacked the beech plot for a short time. The frass-mediated insect faeces appeared to contribute to dissolved and particulate organic matter (Figs. 1 and 2). Here, maximum concentrations of 77.4 mgL−1 (DOC), 108.2 mgL−1 (TOC) and 20.20 mgL−1 (DN) and 22.79 mgL−1 (TN) were observed. May 2007 was characterized by heavy rain events which again occurred subsequently to a drought period before.
Such extreme events occurred less pronounced under spruce, where concentrations were approximately twice as high compared to the beech plot. However, in early April 2006, with notable bark beetle infestation, increased C and N concentrations were recorded.
Correlation analysis (Spearman’s correlation coefficient) between throughfall concentrations of DOC, TOC, DN, TN, NO3-N and solution pH, electrical conductivity (EC), air temperature and throughfall amount of water of the beech and spruce site at Solling (2005–2007).
3.3. Regulating factors for concentration dynamics
Throughfall concentrations were significantly positively correlated with air temperature exhibiting correlation coefficients between 0.30 and 0.68 (Tab. II). Whereas increasing amounts of throughfall water resulted in highly significant negative correlations. Furthermore, the concentrations revealed significant positive relation with solution pH as well as with electric conductivity (Tab. II).
3.4. Matter fluxes with throughfall solution
Although higher throughfall water fluxes were recorded under beech, annual fluxes of C and N fractions were significantly higher under spruce (6–41% for TOC and 9–33% for TN). Consequently, larger fluxes of dissolved and particulate C and N fractions, accumulated over the course of the entire sampling period, were observed in throughfall solution collected under spruce, amounting to 261.8 kgDOCha−1, 312.6 kgTOCha−1, 66.9 kgDNha−1 and 77.7 kg TN ha−1 (Tab. III). Given the fact, that data collection in 2006 at the spruce site was interrupted for approximately 3 months (from August 8th to October 31st) due to bark beetle infestation, accumulated flux amounts are assumed to be even higher compared to those ones under beech.
In average, 80% of DOC and TOC and 70% of DN and TN were released during the growing season at the beech site, compared to 60% for C and N at the spruce site. Slightly lower proportions were observed for NO3-N and residual-N (Tab. IV).
Fluxes of POC at the beech site, relatively stronger contributed to TOC than those at the spruce site (12–19% for spruce and 10–30% for beech, respectively) (Tab. IV). This became particularly apparent during the growing season. Here, 85% of POC were transferred from the canopy to the soil, compared to 65% under spruce (Tab. IV). Fluxes of PON similarly contributed by 10–17% (for spruce) and by 7–20% (for beech), respectively, to the total nitrogen fraction (TN). Proportions of inorganic N species nitrate-N in DN amounted to 42–43% under spruce and to 37–41% under beech, respectively.
The fluxes of dissolved and particulate C and N species were strongly related to the water fluxes exhibiting correlation coefficients (Spearman r) of 0.73 for DOC, 0.75 for POC, 0.65 for DN and 0.63 for PN for spruce and of 0.92 for DOC, 0.89 for POC, 0.75 for DN and 0.72 for PN for beech.
In general, throughfall solutions collected at the beech site revealed significantly (P < 0.05) higher pH values than solutions under spruce, accompanied by significantly lower EC values and C/N ratios in the dissolved and particulate solution fraction (Tab. V). Compared to the dormant season, pH and EC values increased significantly during the growing season, for both tree species (P60.05 for beech and P < 0.001 for spruce).
4. DISCUSSION
Pronounced changes in throughfall composition and element fluxes with time and space have been numerously reported from coniferous and deciduous forests (Currie et al., 1996; de Schrijver et al., 2007; Matzner and Meiwes, 1994). Different interacting processes such as e.g. precipitation patterns (Mercier and Lindow, 2000), tree species canopy architecture (de Schrijver et al., 2007; Levia and Frost, 2006), atmospheric ecosystem deposition (Lamersdorf and Blank, 1995), nutrient leaching from leaf biomass, and immobilization processes within the canopy mediated by phyllosphere micro-organisms (Stadler and Müller, 2000) are under debate to explain the varying energy and nutrient transfer patterns with throughfall solution.
In the present study, we observed tree species related differences in throughfall concentrations and fluxes of dissolved and particulate C and N compounds as well as seasonality effects. In the following we focus on the discussion on seasonality effects encompassing abiotic and biotic factors, tree specific characteristics, canopy architecture, and leaf functionality.
Throughfall solutions do not reflect the element and matter input by bulk precipitation as they underwent chemical alterations during canopy passage, where they interact with various sink and source processes (Draaijers and Erisman, 1995). Consequently, to date it is still difficult to separate internally originating compounds from external inputs. In this context, isotopic labeling appears as a powerful but technically demanding tool for tracking substances in ecosystems. However, the pattern of temporal variations of concentrations and fluxes might provide information with regard to the processes they were subjected to and their abiotic and biotic origin. It is suggested that seasonality-induced patterns of concentrations and fluxes reflect processes of matter accumulation (impaction, interception) on and its release (leaching) from leaves depending on (i) abiotic (physical, meteorological, chemical) and (ii) biotic factors . From this, one can likely conclude on the origin of compounds.
Annual throughfall fluxes (kg ha−1) of DOC, TOC, DN and TN and proportions (%) of DOC and DN in total organic C (TOC) and N (TN).
Annual throughfall fluxes (kg ha−1) of POC, PON, NO3-N and residual-N and proportions (%) of POC and PON in total organic C (TOC) and N (TN).
4.1. Abiotic factors
4.1.1. Water-flux with precipitation
Our results exhibited significant negative correlations between throughfall water-fluxes and C and N concentrations in throughfall solution, which indicate a limited source system, where the pool of releasable matter appeared to be replenished quickly since, throughfall matter fluxes are positively related to water-fluxes. Given the fact that the interception rate of airborne substances remained constant during a season, it is assumable that internal release processes from leaves (accumulation e.g. through excretion) and therefore a contribution to the throughfall appeared temporally (growing season, dormant season) as well as spatially (beech, spruce) dependent. With regard to the precipitation events (duration, intensity and occurrence), internally mediated matter inputs can differ. During periods of low or absent precipitation, compounds may accumulate on leaves and needles (Zhang et al., 2006). With subsequent rainfall compounds are washed-off, causing amplified concentrations and fluxes of C and N compounds with throughfall.
4.1.2. Air temperature
Our results exhibited significant correlations between the C and N concentrations in the throughfall and the air temperature likely promoting the biological production of organic compounds during the growing season. Former studies reported on the correlation between increasing rates of chemical, physical and biological processes within the canopy cover and increased air temperature (Hosker and Lindberg, 1981). The observed seasonal dynamics can be therefore related to more favorable abiotic conditions resulting in an increased release (internal) or deposition (external) of compounds in and on leaf surfaces (Weseley, 1989). For instance, Weseley (1989) reported that the deposition of compounds on leaf surfaces depends on the stomatal resistance, which is conducted by temperature, radiation conditions and vapor pressure deficit.
C/N ratios in dissolved and particulate fractions, electrical conductivity (EC) and pH of the throughfall solutions of the beech and spruce site at Solling (2005–2007).
4.2. Biotic factors
4.2.1. Phyllosphere micro-organisms
With leaf shooting in May (beech) and increasing temperatures, different groups of sessile and mobile organisms in the canopy like yeasts, bacteria, filamentous fungi and (Kinkel, 1997) epiphytic plants become active and thus contribute to matter decay, chemical transformation and nutrient immobilization and release processes into throughfall solutions (Berg and Meentmeyer, 2002). This aspect might explain, why during the growing season with increasing tree physiological activity, enhanced C and N concentration were observed at both forest sites. Furthermore, the enhanced concentrations of the residual-N-fraction (DON + NH4-N) in throughfall might point to organic matter production resulting from microbial activity (Stadler and Müller, 2000). Our results demonstrate that the particulate fraction of C and N compounds occurred mainly during enhanced matter production and release from the leaf shooting period onwards and during the activity of herbivore and xylobiontic insects like in 2006.
Particulate organic carbon considerably adds to DOC by about 30% (beech) and 16% (spruce) over the whole sampling period. Additional amounts by PON revealed values between 14.7% (beech) and 12.1% (spruce). We suggest that the microbial biomass functions as main source for the production of particulate (unfiltered) matter. Our suggestion is based on the fact, that highest concentrations of the particulate fraction appeared in throughfall solution colleted during the growing season compared to the dormant season (beech: +25.5% for POC, +17.9% for PON; spruce: +17.2% for POC; +17.8% for PON), where the biological activity is reduced.
4.2.2. Insect herbivory
How remarkably biological processes can affect throughfall composition is demonstrated by various studies on canopy-active insects and their contribution to altered throughfall solution composition (le Mellec and Michalzik, 2008). Herbivory can alter the throughfall composition by enhancing the organic N and especially the organic C concentrations. This appeared as strong concentration peak in September 9th, where the beech stand was infested by the leaf-feeding lymantrid moth (Dasychira pudibunda). Under infestation and due to leaf feeding activity high amounts of foliar material were transformed into faeces and entered the soil system as solid or particulate C and N compounds.
In 2006 the spruce stand was also infested by the six dentated bark beetle (Pityogenes chalcographus) and the European spruce bark beetle (Ips typograhus). It is suggested, that needle excretion increased due to altered tree physiological circumstances from which enhanced C and N concentrations in the throughfall chemistry derived. The peak in August 2006 firstly indicates this phenomenon.
4.2.3. Canopy properties
Rainfall composition is altered as the water passes the canopy system (Lovett and Lindberg, 1993). The foliage and branches function as major nutrient-specific sinks and sources, consequently retaining and removing nutrients on the sites (Prescott, 2002). The release and absorption (sink/source function) depends on structural properties of the canopy and reactive leaf surfaces, too.
4.2.4. Tree species effect
It is suggested that during tree shooting at broadleaved sites, nutrient up-take from the soil is amplified whereas concurrently the cuticle layer has not been established completely. In particular, the epicuticular wax layer which protects leaves from high water and nutrient losses is absent, from which increased leaching losses might result. Enhanced C and N concentrations at the end of the summer are presumably derived from leaf senescence, when the cuticle layer becomes thinner and permeable due to decomposition processes (Ibrom, 1993).
4.2.5. Canopy structure
Coniferous canopies differ with regard to structural and functional aspects from those of broadleaved ones. This aspect is attested by our results which exhibited higher C and N concentrations in throughfall solution collected under spruce compared to those of beech, especially during the growing season when both stands are foliated. Our findings corroborate the results observed in other studies which report on higher throughfall depositions in coniferous stands (de Schrijver et al., 2007). The canopy structure influences the aerodynamic resistance and deposition velocity, on which the airborne accumulation depends. Furthermore, the deposition speed depends on the horizontal wind velocity and the structure of the acceptor surface. The structure of spruce twigs including their needles favors the deposition by impaction and diffusion processes. In this context, coniferous canopies show a higher filtering capacity to airborne compounds than those of broadleaved ones exhibiting a higher interception rate of 30 to 40% of the gross precipitation, while beech canopies only intercept 17–20% of the gross precipitation (Rothe et al., 2002). Furthermore, tree habits and architecture of beech and spruce differ in such manner, that in beech stands water fluxes with nutrients are guided into the tree center (stem), resulting in high stem flows (up to 20% of the total precipitation) (Chang and Matzner, 2000), whereas water fluxes in spruce stands are guided away from the canopy center resulting in higher fluxes from the overall canopy (Seiler and Matzner, 1995).
Higher concentrations of C and N compounds in throughfall solutions at the spruce site might be explained by internal and external inputs. Interception at spruce trees is higher resulting in higher accumulation rates of airborne particles, due to a denser foliage, a higher LAI (Leaf Area Index) of 20.5 compared to 6.5 m2m−2 at the beech site, higher needle surface roughness and higher leaf longevity (Rothe et al., 2002).
5. CONCLUSIONS
The results of our field experiment clearly support the hypothesis that the canopy is a relevant source for particulate organic matter (POM) in forest ecosystems, representing between 20–30% of the TOC and 10–20% of the TN fluxes with throughfall. We further found, that the quantity and temporal dynamics of DOM and POM in broadleaved forests differ from those in coniferous ones, in that element concentrations were twice as high and throughfall fluxes at least 1.6 times higher under spruce than under beech. The canopy-derived POM is built up by different sources, which we found to encompass pollen deposition, excretions of phytophagous insects, atmospheric deposits and temperature driven microbial-derived products. Environmental factors such as air temperature and rain patterns (dry and wet periods) promote the production and release of POM.
Acknowledgments
We are deeply indebted to Petra Voigt and Anja Soedje from the Laboratory Unit of the Institute of Geography at the University of Göttingen for chemical analyses and help during the study and to Annett Reinhardt and Björn Geschwind for their great help in the field. Many thanks also to Andreas Schulze of the NW-FVA (Nordwestdeutsche Forstliche Versuchsanstalt) for helping with the ECO data base at NW-FVA. Financial support was provided by the German Research Foundation (DFG, Project No. MI 927/1-3) and the German Ministry for Research and Technology (BMBF) supporting this research within the Juniorprofessur programme.
References
- Aber J.D., Nadelhoffer K.J., Steudler P., and Melillo J.M., 1989. Nitrogen satauration in northern forest ecosystems: hypotheses and implications. BioScience 39: 378–386. [CrossRef] [Google Scholar]
- Berg B., and Meentemeyer V., 2002. Litter quality in a north European transect versus carbon storage potential. Plant Soil 242: 83–92. [CrossRef] [Google Scholar]
- Carlisle A., Brown A.H.F., and White E.J., 1966. Litter fall, leaf production and the effects of defoliation by Tortrix viridana in a sessile oak (Quercus petraea) woodland. J. Ecol. 54: 65–85. [CrossRef] [Google Scholar]
- Chang S.-C., and Matzner E., 2000. The effect of beech stemflow on spatial patterns of soil solution chemistry and seepage fluxes in a mixed beech/oak stand. Hydrol. Process. 14: 135–144. [CrossRef] [Google Scholar]
- Currie W.S., Aber J.D., McDowell W.H., Boone R.D., and Magill A.H, 1996. Vertical transport of dissolved organic C and N under long-term N amendments in pine and hardwood forests. Biogeochemistry 35: 471–505. [CrossRef] [Google Scholar]
- De Schrijver A.D., Geudens G., Augusto L., Staelens J., Mertens J., Wuyts K., Gielis L., and Verheyen K., 2007. The effect of forest type on throughfall deposition and seepage flux: a review. Oecologia 153: 663–674. [CrossRef] [PubMed] [Google Scholar]
- Draaijers G.P.J., and Erisman J.W., 1995. A canopy budget model to assess atmospheric deposition from throughfall measurements. Water Air Soil Pollut. 85: 2253–2258. [CrossRef] [Google Scholar]
- Ellenberg H., Mayer R., and Schauermann J., 1986. Ökosystem-forschung- Ergebnisse des Sollingprojektes. Ulmer Verlag, Stuttgart. [Google Scholar]
- Finzi A.C., Canham C.D., and v. Breemen N., 1998. Canopy tree-soil interactions within temperate forests. Ecol. Appl. 8: 447–454. [Google Scholar]
- Hagedorn F., van Hees P.A.W., Handa, I.T., and Hättenschwiler, S., 2008. Elevated atmospheric CO2 fuels leaching of old dissolved organic matter at the alpine treeline. Global Biogeochem. Cycles 22. DOI: 10.1029/2007GB003026. [Google Scholar]
- Hosker R.P. Jr., and Lindberg S.E., 1981. Review: atmospheric deposition and plant assimilation of gases and particles. Atmosph. Environ. 16: 889–910. [CrossRef] [Google Scholar]
- Ibrom A. 1993. Die Deposition und die Pflanzenauswaschung (Leaching) von Pflanzennährstoffen in einem Fichtenbestand im Solling, Forschungszentrum Waldökosysteme, Reihe A, Bd. 105. [Google Scholar]
- Kinkel L.L., 1997. Microbial population dynamics on leaves. Annu. Rev. Phytopathol. 35: 327–347. [CrossRef] [PubMed] [Google Scholar]
- Lamersdorf N.P. and Blank K., 1995. Evaluation of fine material input with throughfall for a spruce forest in Solling, FRG, by means of a roof construction. In: Jenkins A., Ferrier R.C., Kirby C. (Eds.), Ecosystem manipulation experiments: scientific approaches, experimental design and relevant results, Ecosystem Research Report 20, Commission of the European Communities. [Google Scholar]
- Le Mellec A., and Michalzik B., 2008. Impact of a pine lappet (Dendrolimus pini) mass outbreak on C and N fluxes to the forest floor and soil microbial properties in a Scots pine forest in Germany. Can. J. For. Res. 38: 1829–1849. [CrossRef] [Google Scholar]
- Levia Jr., D.F., and Frost E.E., 2006. Variability of throughfall volume and solute inputs in wooded ecosystems. Progr. Phys. Geogr. 30: 605–632. [CrossRef] [Google Scholar]
- Lichter J., Lavine M., Mace K.A., Richter D.D., and Schlesinger W.H., 2000. Throughfall chemistry in a loblolly pine plantation under elevated atmospheric CO2 concentrations. Biogeochemistry 50: 73–93. [CrossRef] [Google Scholar]
- Lovett G.M., and Lindberg S.E., 1993. Atmospheric deposition and canopy interactions of nitrogen in forests. Can. J. For Res. 23: 1603–1616. [CrossRef] [Google Scholar]
- Manderscheid B., and Matzner E., 1995. Spatial heterogeneity of soil solution chemistry in a mature Norway spruce (Picea abies (L.) Karst.) stand. Water Air Soil Pollut. 85: 1185–1190. [CrossRef] [Google Scholar]
- Matzner E., and Meiwes K.J., 1994. Long-term development of element fluxes with bulk precipitation and throughfall in two German forests. J. Environ. Qual. 23: 162–166. [CrossRef] [Google Scholar]
- Meesenburg H. and Brumme R., 2009. General Description of the Study site. In: Functioning and Management of European Beech Ecosystems, Ecological Studies 2008, Springer Verlag, Berlin, Heidelberg. [Google Scholar]
- Mercier J., and Lindow S.E., 2000. Role of leaf surface sugars in colonization of plants by bacterial epiphytes. Appl. Environ. Microbiol. 66: 369–374. [CrossRef] [PubMed] [Google Scholar]
- Michalzik M., Kalbitz K., Park J.H., Solinger S., and Matzner E., 2001. Fluxes and concentrations of dissolved organic carbon and nitrogen – a synthesis for temperate forests. Biogeochemistry 52: 173–205. [CrossRef] [Google Scholar]
- Panferov O., Kreilein H., Meesenburg H., Eichhorn J., and Gravenhorst G., 2010. Climate at three beech forest sites in Central Germany, In: Brumme R. and Alphei J. (Eds.), Human impacts on carbon and nitrogen cycles in temperate beech forests, Ecological Series, Springer, Berlin/New York, Germany (in print). [Google Scholar]
- Prescott C.E., 2002. The influence of forest canopy on nutrient cycling. Tree Physiol. 22: 1193–1200. [PubMed] [Google Scholar]
- Rennenberg H., and Gessler A., 1999. Consequences of N deposition to forest ecosystems – recent results and future reserach needs. Water Air Soil Pollut. 116: 47–64. [CrossRef] [Google Scholar]
- Rothe A., Huber C., Kreutzer K., and Weis W., 2002. Deposition and soil leaching in stands of Norway spruce and European Beech: results from the Höglwald research in comparison with other European case studies. Plant Soil 240: 33–45. [CrossRef] [Google Scholar]
- Schipka F., Heimann J., and Leuschner C., 2005. Regional variation in canopy transpiration of Central European beech forests. Oecologia 143: 260–270. [PubMed] [Google Scholar]
- Seiler J., and Matzner E., 1995. Spatial variability of throughfall chemistry and selected soil properties as influenced by stem distance in a mature Norway spruce (Picea abies (L.) Karst.) stand. Plant Soil 176: 139–147. [CrossRef] [Google Scholar]
- Stadler B., and Müller T., 2000. Effects of aphids and moth caterpillars on epiphytic micro-organisms in canopies of forest trees. Can. J. For. Res. 30: 631–638. [CrossRef] [Google Scholar]
- Sollins P., Grier C.C., McCorison F.M., Cromack Jr. K., Fogel R., and Fredriksen R.L., 1980. The internal element cycles of an old-growth Douglas-fir ecosystem in Western Oregon. Ecol. Monogr. 50: 261–285. [CrossRef] [Google Scholar]
- Weseley M.L., 1989. Parameterization of surface resistances to gaseous dry deposition in regional-scale numerical models. Atmos. Environ. 23: 1293–1304. [CrossRef] [Google Scholar]
- Watanabe, M., Takamatsu T., Koshikawa M.K., Yamamura S., and Inubushi K., 2008. Dry deposition of acidic air pollutants to tree leaves, determined by a modified leaf-washing technique. Atmos. Environ. 42: 7339–7347. [CrossRef] [Google Scholar]
- Wright R.F., Alewell C., Cullen J.M., Evans C.D., Marchetto A., Moldan A., Prechtel A., and Rogora M., 2001. Trends in nitrogen deposition and leaching in acid-sensitive strams in Europe. Hydrol. Earth Syst. Sci. 5: 299–310. [CrossRef] [Google Scholar]
- Zhang G., Zeng G.M., Jiang Y.M., Huang G.H., Li J.B., Yao J.M., Tan W., Xiang R.J., and Zhang X.L., 2006. Modelling and measurement of two layer-canopy interception losses in a subtropical evergreen forest of south-central China. Hydrol. Earth Syst. Sci. 10: 65–77. [CrossRef] [Google Scholar]
All Tables
Average annual concentrations (mg L−1) of different C and N fractions in throughfall solutions (mean ± standard deviation) of the beech and spruce site at Solling (2005–2007).
Correlation analysis (Spearman’s correlation coefficient) between throughfall concentrations of DOC, TOC, DN, TN, NO3-N and solution pH, electrical conductivity (EC), air temperature and throughfall amount of water of the beech and spruce site at Solling (2005–2007).
Annual throughfall fluxes (kg ha−1) of DOC, TOC, DN and TN and proportions (%) of DOC and DN in total organic C (TOC) and N (TN).
Annual throughfall fluxes (kg ha−1) of POC, PON, NO3-N and residual-N and proportions (%) of POC and PON in total organic C (TOC) and N (TN).
C/N ratios in dissolved and particulate fractions, electrical conductivity (EC) and pH of the throughfall solutions of the beech and spruce site at Solling (2005–2007).
All Figures
|
Figure 1 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved and particulate organic C fractions in throughfall solutions of the beech stand (2005–2007). |
| In the text | |
|
Figure 2 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved and particulate organic C fractions in throughfall solutions of the spruce stand (2005–2007). |
| In the text | |
|
Figure 3 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved and particulate N fractions in throughfall solutions of the beech stand (2005–2007). |
| In the text | |
|
Figure 4 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved and particulate N fractions in throughfall solutions of the spruce stand (2005–2007). |
| In the text | |
|
Figure 5 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved N fractions in throughfall solutions of the beech stand (2005–2007). |
| In the text | |
|
Figure 6 Weekly (2005) and fortnightly mean concentrations (mgL−1) and standard deviations (SD) of dissolved N fractions in throughfall solutions of the spruce stand (2005–2007). |
| In the text | |